
蝙蝠(学名:Chiroptera)是脊索动物门、哺乳纲下的一类动物,是唯一能够真正飞翔的哺乳动物;除一般哺乳动物的特点外,还有一系列适应飞行的形态特征 。最小的是混合蝠,体重仅有1.9克,翼展16厘米;一些狐蝠的体重可以超过1.3千克,翼展可达1.7米。蝙蝠全身骨质轻,头骨愈合程度较高,肩带发达,胸骨具龙骨状突起。后肢胫骨、腓骨退化。耳壳发达,常具发达的耳屏或对耳屏。翼膜、耳、唇等处有丰富的感觉毛。颜色多样,大多是褐色、灰色和黑色。
蝙蝠具有很强的飞行能力,同时也是多种人畜共患病毒的天然宿主,能够携带数十种病毒 。蝙蝠与其能够飞行并进行夜间生活相适应,它们在生理机能上也发生了一系列重要变化。通常蝙蝠的视觉较差,而听觉则异常发达,在夜间或十分昏暗的环境中它们能够自由地飞翔和准确无误地捕捉食物 。70%的蝙蝠种类主要以昆虫和其他小节肢动物为食;其余的种类主要以果实、花蜜和花粉为食;一些热带种类是食肉的,有3个种类的吸血蝙蝠以血液为食 。分布于地球上除南北极和某些大洋岛屿以外的所有地方 。
蝙蝠可分为两个亚目,全世界共有16科185属962种,中国约7科30属120种。1996年世界自然保护联盟物种生存委员会(IUCN/SSC)出版的《1996年受胁动物红色名录》中公布了蝙蝠(翼手目)动物受胁状况,指出处于极危险(CR)种类有26种,濒危(EN)有32种,易危(VN)有173种。许多蝙蝠在自然界越来越少,趋于灭绝,严重的物种濒危局面令人深思
- 蝙蝠
- Chiroptera
- 天鼠、挂鼠、天蝠、老鼠皮翼、飞鼠、燕别故、蜜符、岩老鼠
- 动物界
- 脊索动物门
- 脊椎动物亚门
- 哺乳纲
物种进化
已知最早的蝙蝠化石是有5000万年历史的食指伊神蝠,是在美国怀俄明州发现的。后来发现德国的麦塞尔湖湖床层内有更完整的蝙蝠化石,这些化石可以追溯到始新世时期,它们的胃里包含有成了化石的昆虫。这些早期的蝙蝠有许多和现存的小蝙蝠亚目相似,因而很难重现它们进化中的早期阶段,但是它们耳蜗(内耳)的结构表明它们几乎肯定是使用回声定位的。最早的与大蝙蝠亚目相似的化石,可追溯到大约3500万年前。
有些科学家提出,与小型蝙蝠相比,大型蝙蝠可能实际上与灵长类的关系更近。除了没有回声定位系统之外,大型蝙蝠和灵长类都有高级的视觉能力,而且它们指骨的长度也有相似的比例。科学家推测,翼手目两个亚目里是分别独立进化出来的,然而这一理论却无法得到分子学研究的证实。相反分子学研究证明了蝙蝠(翼手目)群体内的相似性,因为两个亚目的基因里都含有较高水平的腺嘌岭和胸腺嘧啶碱基对。支持“飞行的灵长类”观点的科学家反对上述观点,他们说基因的相似性可能仅仅是因为两个亚目在飞行中都需要消耗大量的能量,是进化动力在相似的条件下将它们独自塑造而成,而不是因为它们有一个近期的共同祖先。大部分分子学研究现在支持蝙蝠(翼手目)的单一起源理论,即狐蝠和小型蝙蝠有共同的祖先。
分子学证据还表明,各种菊头蝠和它们的近亲诸如旧大陆假吸血蝠,与大蝙蝠亚目的亲缘关系要比和其他小型蝙蝠更近。然而让人疑感的是,菊头蝠拥有一些动物界里最复杂的回声定位系统,而大型蝙蝠几乎没有。这种理论意味着要么是菊头蝠独立于其他小型蝙蝠进化出了回声定位系统,要么就是大型蝙蝠曾经拥有类似的能力只是后来丢失了。以上两种理论任何一个似乎都有可能。第三种可能性是这种分子学证据其本身可能是误导。
分类研究
基因研究
中外科学家完成的蝙蝠基因组学研究在《科学》上在线发表。科学家对两种不同类群的蝙蝠基因组比较分析,揭示了蝙蝠飞行及免疫系统的适应性相关机制,阐明不同蝙蝠类群的分子多样性机制,为蝙蝠及其它哺乳动物在生物学及进化等方面的研究提供了新思路。
蝙蝠具有很强的飞行能力,同时也是多种人畜共患病毒的天然宿主,能够携带数十种病毒,还是唯一演化出具有真正飞行能力的哺乳动物,其飞行能力的进化与一系列复杂的形态和生理变化息息相关。通过深入分析,研究人员发现一系列与DNA损伤检验点或DNA修复通路相关的基因在蝙蝠中受到了很强的正选择作用。他们还发现与皮肤弹性相关的基因和参与肌肉收缩的基因在蝙蝠中发生了快速进化,可能也有助于飞行。
蝙蝠可携带多种人畜共患病毒,自身几乎不受感染。通过对相关基因的研究,科学家发现蝙蝠中NF-κB家族转录因子c-REL受到正选择。该基因不仅在固有免疫中发挥功能,还与DNA损伤反应具有一定关系。自然杀伤性(NK)细胞是抵抗外界病原微生物和肿瘤的第一道防线。
研究人员表示,基因组学的发展为人类了解物种起源、分化、多样性的遗传基础提供了重要的基础数据。蝙蝠在进化中具有特殊的地位,在长期演化过程中发展出许多非常有趣的生物学现象,比如飞行、回声定位、冬眠等。基因组学只是开展这些研究的入口之一,研究的数据和结果对相关研究具有重要意义。同时,蝙蝠是对人类具有极大危害的病毒载体,基因组学层面的比较分析,也将为了解蝙蝠自身的免疫系统和病毒防卫机制提供重要的工具。
该成果由华大基因、澳大利亚动物研究所、中科院武汉病毒所、美国海军医学中心及亨利·杰克逊基金会等机构的科学家共同完成。
形态特征
共同特征
蝙蝠前肢特化,尤其是骨骼有较大的变化。肱骨显著短于桡骨(前臂骨),尺骨退化;除第一指不特别延长,末端有爪,其余掌骨和指骨均特别延长,各掌、指骨间生有皮膜,向后一直与后肢和尾部相连。连接各指间的皮膜称翼膜,前肢肱骨和后肢间皮膜称侧膜,前肢肱骨和前臂骨前的皮膜称前翼膜,连接左右后肢和尾部的称股间膜。股间膜其膜缘常有距,起支撑股间膜作用,距外侧皮膜称之为距缘膜。后肢短小,大腿部与身体呈直角且位于同一平面。侧膜常止于胫下部、踵部或趾基部。后肢足部完全位于皮膜以外,五趾均具爪,用于钩挂。
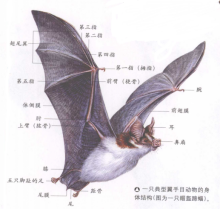
分类特征
- 头骨与牙齿
蝙蝠的头骨和牙齿的形状主要和所吃的食物有关。杂食的叶口蝠和食虫的圆耳蝠(四耳蝠属)有最普通的牙齿以便嚼碎食物,而大食果蝠有大而平的臼齿。吸食花蜜的无尾长舌蝠很少使用牙齿,然而它们的头骨被拉长以容下一条长而多毛的舌头。普通吸血蝠凿子一样的门齿和竖向扁平像剃刀一样的上犬齿被用来在动物皮肤上咬开一个槽形切口,以便于它在里面舔食血液。肉食的假吸血蝠有巨大的头骨,强壮尖锐的犬齿和剪刀一样的日齿,可以削骨切肉。
- 翅膀
蝙蝠的翅膀呈现两个典型的类型,反映了两种不同的比例,即翼展和翼宽的比例。有些种类诸如大菊头蝠(a处),这一比例比较小,它能适应在树叶之间低速而精巧地飞行其他种类诸如墨西哥皱唇蝠(b处),这一比例比较大,能适应没有障碍物的高速飞行。
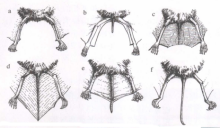
- 尾部
不同属的蝙蝠在尾部形状上反映出主要的不同:a尾巴完全缺失(狐蝠属);b短尾(管鼻果蝠属);c鞘尾(鞘尾蝠属);d全膜(鼠耳蝠属);e散尾(皱唇蝠属);f鼠尾(鼠尾蝠属)。
生活习性
活动特性
蝙蝠的体温变幅很大,可达56℃(由-7.5-48.5℃),某些大型的食果蝠,由于其生理和行为特点不同,体温比较恒定,而绝大多数的小型食虫蝠则是典型的异温动物。据观察,生活在温带的蝙蝠,一生中其不活动的时间比活动的时间要多得多,夏天它们差不多整个白天和夜间的一部分时间都在睡眠,有些种类当天气寒冷的时候,便像鸟类一样进行长距离的季节性迁徒,到南方去过冬,留在原地的种类则在严寒的冬季便进入长时期的冬眠。据不完全统计,栖息在列宁格勒附近的大耳蝠和北棕蝠在一生中其活动的时间仅占1/15~1/20,其余的时间都在睡眠(库加金,1959)。当环境温度达到16~28℃时,蝙蝠即处于深睡状态,睡眠的蝙蝠生理机能减慢,新陈代谢降低。由于代谢低耗,致使某些蝙蝠寿命较长。
回声定位
蝙蝠与其能够飞行并进行夜间生活相适应,它们在生理机能上也发生了一系列重要变化。通常蝙蝠的视觉较差,而听觉则异常发达,在夜间或十分昏暗的环境中,它们能够自由地飞翔和准确无误地捕捉食物,最基本的手段是能够利用回声定位。实验证明,多数蝙蝠是利用从喉头发出的超声脉冲来定位的。但也不尽相同,某些大型的食果蝠如棕果蝠,其回声定位的能力比较特殊,它们是利用咂舌的发声作为声音定位依据的。回声定位机能对于蝙蝠的生活来说是十分重要的,使其能够在夜间或较为昏暗的环境中占据鸟类食虫无法利用的生态位,而这些地方在白天却是各种鸟类的生活领城。

多数蝙蝠叫声的频率在20~60千赫之间。频率低于20千赫的声波波长大于多数昆虫声波的波长,因此会穿过昆虫而不会反射回来;频率高于60千赫时在空气中衰减得很快,这就限制了其可使用的范围,因此多数蝙蝠叫声的频率不会高于60千赫。但一些飞行速度比较慢的蝙蝠却可以对高频率的声波做出较为轻易的反应,高频率的声波是它们理想的应用“武器”。短耳三叶鼻蝠能发出特高频率的叫声,其频率可达212千赫,是所有蝙蝠中最高的;而有些犬吻蝠科的蝙蝠,例如小斑点蝠回声定位的频率却可以低到11千赫。运用特别高或特别低频率叫声的另外一个好处就是这些频率的声波很难被捕食的猎物一昆虫感知到,这样蝙蝠就可以捕捉更多的昆虫。
食性特点
社交能力
蝙蝠也会交谈。一些社交性的信号可以用来吸引配偶、保卫食物、召唤同类以及驱逐一些掠食蝙蝠的动物。这些信号通常采用很低的频率发出(有的时候人类也可以听到),因此,这些声音可以传播很远。同时,这些声音也会因为发声蝙蝠的不同而不同,比如幼小的蝙蝠在和母蝙蝠分开以后会发出“孤独信号”,母蝙蝠会根据发声的不同迅速地分辨出是否是自己的幼崽。另外,许多小型蝙蝠对它们经常使用的回声的频率也很敏感。
分布范围
繁殖方式
整个蝙蝠群的性周期是同步的,因此大部分交配活动发生于数周之内。孕期多不相同,在延迟着床的情况下同一种类的时间差别为3~10个月。许多种类的雌体妊娠后迁到一个特别的哺育栖息地点。蝙蝠每年繁殖一次,每产1~2崽;幼仔初生时无毛或少毛,常在一段时间内不能视不能听;幼崽生长很快,一般在6~8周龄时即可达到成体大小,1~2月龄即脱去胎毛。幼仔由亲体照顾5周~5个月,按不同种类决定。寿命最长可达33年,大多数种类可能平均4~5年。
下级分类
翼手目可以分为两个亚目:大蝙蝠亚目(食果蝠)和小蝙蝠亚目(食虫蝠)。蝙蝠类动物全世界共有16科185属962种,中国约有7科30属120种。
中文名 |
拉丁学名 |
简要介绍 |
|---|---|---|
大蝙蝠亚目 |
Megachiroptera |
体型一般较大。眼大,视觉好,能在弱光下不依赖回声定位飞行。吻较长。以花果为食。仅1科。 |
狐蝠科 |
Pteropodidae |
有42属166种,分为2个亚科:狐蝠亚科(Pteropodinae)和长舌狐蝠亚科(Macroglossinae)。分布于旧大陆热带、亚热带地区,以东南亚和非洲种类最多。中国有5属7种,见于华南区,包括台湾和海南省。均为植食性,其中大型的种类多以果实为食,小型种类主要食花蜜。夜行性远距离飞行觅食,有时可达15公里。主要靠嗅觉发现食物。仅棕果蝠等少数属有超声定位功能。大型者多聚居,小型者多独栖。终年繁殖,或集中在9-11月间,翌年2月产仔。最多每年1胎,每胎1-2仔。饲养条件下可活20年。 |
小蝙蝠亚目 |
Microchiroptera |
多数体型较小。眼小,回声定位系统发达。吻较短。以食昆虫为主,少数食花果或吸血。包括15科。 |
鼠尾蝠科 |
Rhinopomatidae |
有1属3种。分布于亚洲南部到非洲北部。无鼻叶的中小型的蝙蝠。最显著的特征是有非常长的尾巴,是所有蝙蝠中尾巴最长者,几乎和身体等长。食虫性。比较适应干旱和半干旱的环境。 |
凹脸蝠科(混合蝠科) |
Craseonycteridae |
仅1属1种,即凹脸蝠(混合蝠)(Craseonycteris thonglongyai)。于1973年发现于泰国西部。体长仅3厘米左右,体重不到2克,是最小的翼手目动物,但翼展却有17厘米。口鼻部略似猪鼻,无鼻叶,耳大而有隆起的耳屏,无尾。以昆虫为食。 |
鞘尾蝠科 |
Emballonuridae |
有13属46种。广布于世界热带、亚热带地区。无鼻叶,尾部被尾膜包裹而尾尖突出在尾膜上,故名。喉部有可以分泌芳香分泌物的小囊,用以吸引异性。适应多种不同生活环境,但是一般不远离树木,主要食昆虫,有时也食果实作为补充。 |
假吸血蝠科(巨耳蝠科) |
Megadermatidae |
仅4属5种。分布于非洲至大洋洲。体大,无尾,鼻叶发达,耳大而圆。原认为吸血,因而引人注意,现知其食肉,以其他蝙蝠、蜥蜴、小鼠等为食,故名。 |
菊头蝠科 |
Rhinolophidae |
有10属129种,包括2亚科:菊头蝠亚科(Rhinolophinae)(1属63种)和蹄蝠亚科(Hipposiderinae)(9属66种)。主要分布于世界热带和亚热带地区,少数分布于温带。因有结构比较复杂的马蹄形鼻叶而得名,蹄蝠类稍弱,从鼻孔而非从嘴中发出声纳,耳朵较大但没有耳屏。常不同种十几只或几十只集群共居一个山洞,季节性出现同性群。不畏光,不甚畏人。捕食蚊、鳞翅目昆虫。 |
兔唇蝠科 |
Nocilionidae |
仅1属2种。分布于拉丁美洲的。体强壮。兔唇蝠Noctilio leporinus又称猛犬蝠,食鱼蝠,以食鱼著称,是仅有的以鱼为主食的蝙蝠。口鼻部尖而没有鼻叶,耳朵大而有小的耳屏,爪子锋利,可以抓住长达10厘米的小鱼,也能用爪子捕捉昆虫。 |
髯蝠科(妖面蝠科) |
Mormoopidae |
有2属8种。分布于美国亚利桑那州到巴西之间。无鼻叶,但唇和颊有复杂的皮褶,下唇似盘状膨大,耳很大并有复杂的耳屏。食虫性,其嘴部结构可能有助于捕食昆虫,喜群居,常在洞穴中结成大群。包括妖面蝠属(怪脸蝠属)(Mormoops)和裸背蝠属(Pteronotus)。 |
叶口蝠科 |
Phyllostomatidae |
有50属139种,分为8个亚科:叶口蝠亚科(Phyllostomatinae)、长舌蝠亚科(Lonchophyllinae)、狭叶蝠亚科(Brachyphyllinae)、花叶蝠亚科(Phyllonycterinae)、长舌叶口蝠亚科(Glossphaginae)、短尾叶口蝠亚科(Garolliinae)、尖皮蝠亚科(Stenodermatinae)、吸血蝠亚科(Desmodontinae)。分布于南美洲。鼻叶发达,有耳屏。体型和习性多样化。食性复杂,包括食虫、食果实或花蜜、食肉甚至吸血者。吸血蝠亚科的3种吸血蝠有时也单列为吸血蝠科Desmodontidae,是陆生脊椎动物中仅有的吸血的成员,常在地面用四肢灵活行走。 |
长腿蝠科(筒耳蝠科) |
Natalidae |
仅1属5种。分布于南美洲。体型较小,体长仅3~5.5厘米。两鼻孔接近,无鼻叶;耳朵大而呈漏斗型,有厚而弯曲的耳屏。四肢和尾均细长,尾被尾膜所包裹。雄蝠额头上有一个鳞茎状的分泌腺,其功能不明。食虫性,居住在多岩洞地区。 |
上一篇:大狐蝠
下一篇:无
|
